АВЛеЪЁЪГгУОњММЪѕаЛс
ФњЕФЮЛжУЃКЪзвГ > ЪГгУОњбЖЯЂ
ЕШРызгЬхгеБфСщжЅММЪѕМАЙтЦзЩИбЁЗНЗЈбаОП
ЗЂВМШеЦкЃК2019/11/1 11:59:45
ЛЦЧр1ЃЌ*ЃЌТэгёК1,2ЃЌвІЙњЛЊ1, КЮЛЊЦц2, еХйЛйЛ1
1.жаЙњПЦбЇдККЯЗЪЮяжЪПЦбЇбаОПдК
2.АВЛеПЦММбЇдК
*ЭЈбЖзїепЃКhuangq@ipp.ac.cn
еЊвЊЃКзїЮЊИЛКЌЖржжвЉРэГЩЗжЕФвЉгУОњМАДЋЭГжавЉВФЃЌСщжЅздЙХвдРДОЭЪмЕНЙуЗКЕФжиЪгЁЃЮЊСЫНјвЛВНЬсИпЯжгаГрСщжЅЕФгааЇЛюадГЩЗжЃЌПЩвдЖдЦфНјаагеБфбЁг§ЁЃЕЭЮТЕШРызгЬхзїЮЊвЛжжаТаЭИпаЇЕФгеБфЪжЖЮЃЌдкСщжЅг§жжжаЕУЕНСЫгІгУ,гЩДЫПЩвдЛёЕУЗсИЛЕФЭЛБфжжжЪзЪдДЁЃСэвЛЗНУцЃЌдкСщжЅг§жжбаОПжаЛЙашвЊЗЂеЙЖдЭЛБфОњжъПьЫйгааЇМјБ№ЗжЮіМАЩИбЁЕФЗНЗЈЃЌетвВЪЧФПЧАбаОПЕФШШЕуКЭФбЕуЁЃБОЮФНщЩмСЫНќФъРДЮвУЧгІгУЕЭЮТЕШРызгММЪѕНјааСщжЅгеБфг§жжЗНУцЕФЙЄзїЃЌвдМАЙигкгІгУКьЭтЙтЦзЗНЗЈЖдгеБфСщжЅЦЗжжНјааЦЗжЪЗжЮіМАЙтЦзЩИбЁЗНЗЈЕФЬНЫїЁЃ
вЛЁЂЧАбд
СщжЅЪЧвЛжжвЉгУецОњЃЌОпгаВЙЦјАВЩёЁЂЗіе§ЙЬБОЕШЙІаЇЁЃЫќЪЧЮвЙњДЋЭГжавЉЃЌРњЪЗгЦОУЃЌВЂвбЪеТМгкЖрИіЙњМвЕФЯжДњвНбЇвЉЕфЁЃвЛАуШЯЮЊГрСщжЅЕФЖрЬЧКЭШ§нЦЪЧЦфжївЊЕФвЉгУГЩЗжЁЃвђДЫЃЌШчЙћФмЬсИпСщжЅЖрЬЧКЭСщжЅШ§нЦЕФКЌСПЃЌНЋЖдСщжЅВњвЕЗЂеЙгаживЊвтвхЁЃЕЋЪЧвАЩњСщжЅОњжжзЪдДгаЯоЃЌашвЊНјвЛВННјааОњжъбЁг§КЭШЫЙЄддХрЃЌЫљвдЙЙНЈДѓСПЭЛБфЬхПтВЂХфКЯвдДѓЙцФЃЩИбЁДыЪЉЪЦдкБиааЁЃФПЧАЃЌвбгааэЖрбЇепПЊеЙетЗНУцЕФбаОПЃЌШчPeng ЕШгУТШЛЏяЎКЭTriton X-100геБфСщжЅдЩњжЪЬхЃЌЭЈЙ§ЩИбЁЕУЕНЕФЭЛБфЬхжъЃЌЦфАћФкЖрЬЧКЭСщжЅШ§нЦКЌСПЗжБ№ЬсИпСЫ568.45%КЭ373.43%[1]ЁЃYeongЕШЖдЩёжл1КХЩЯДюдиОЬЋПегеБфЕФСщжЅОњжъВЩгУЙтЦзЗНЗЈНјааЗжЮі[2]ЁЃЕЫДКУЗЕШЖдЮвЙњЗЕЛиЪНЮРаЧЩЯДюдиЕФСщжЅОњжжНјааСЫддХрЪЕбщЃЌЗЂЯжЦфжаЕФЮРаЧСщжЅ2КХдкОњЫПЩњГЄЫйЖШКЭОњЫПЬхжаАБЛљЫсКЌСПЁЂзгЪЕЬхВњСПКЭжЪСПЕШЖрИіжИБъЩЯОљгаЫљЬсИп[3]ЁЃеХвјЦМЕШРћгУзЯЭтЯпКЭNTGИДКЯгеБфСщжЅдЩњжЪЬхЃЌбЁг§ЛёЕУУШЗЂЫйЖШПьЁЂГЄЪЦЭњЪЂЕФОњжъ[4]ЁЃЖгёчтЕШРћгУТШЛЏяЎЁЂEMSКЭбЧЯѕЫсбЮЖдСщжЅдЩњжЪЬхНјаагеБфЃЌВЂЭЈЙ§ЩИбЁЕУЕНСЫСщжЅЖрЬЧКЌСПИпЕФОњжъ[5]ЁЃWeiЕШЖдЮЂЩњЮяОњжжПтЕФСщжЅОњжъНјааБШНЯЃЌЩИбЁЕУЕНЗЂНЭЬѕМўЯТЕФСщжЅЖрЬЧКЭШ§нЦКЌСПИпЕФОњжъ[6]ЁЃзюНќЃЌHaoЕШЖдСщжЅНјааСЫCRISPR-Cas9ЛљвђБрМВйзїЃЌЪЕЯжСЫЖдСщжЅжаura3БъМЧЛљвђЕФЧУГ§[7]ЁЃЕЋЪЧЃЌЫћУЧбаОПЗЂЯжЃЌЖдгкЦфЫќШБЗІбЁдёадБэаЭЛљвђЕФЧУГ§ВйзїШДгаРЇФбЃЌЦфжаРЇФбжЎвЛЪЧШБЗІБъМЧЛљвђДгЖјжЦдМСЫЛљвђБрМКѓЕФОњжъЕФЩИбЁЁЃЫљвдЃЌЖдгкСщжЅЕШИпЕШецОњЖјбдЃЌИпаЇЕФгеБфЪжЖЮХфКЯгааЇЕФЩИбЁЗНЗЈЪЧг§жжГЩЙІЕФЙиМќЁЃ
ЖўЁЂЕШРызгЬхгеБфММЪѕдкЮЂЩњЮяг§жжжаЕФгІгУ
ЫфШЛЛљгкдЄЯШЩшМЦЕФЛљвђЙЄГЬЗНЗЈвбГЩЮЊФГаЉЮЂЩњЮяОњжжИФСМЕФЪзвЊЪжЖЮЃЌЕЋдкаэЖрЧщПіЯТЃЌЛљгкЮяРэ/ЛЏбЇгеБфММЪѕЕФЫцЛњгеБфЪжЖЮШдШЛЪЧЪЎЗжживЊЕФЃЌЬиБ№ЪЧдкФГаЉЛљвђВйзїЪжЖЮДцдкММЪѕЛђЗЧММЪѕРЇФбЕФЧщПіЯТЃЌетжжгеБфЗНЗЈОЭЛсеЙЪОГіЦфЬигаЕФгХЪЦЁЃЯрБШгкЦфЫќЮяРэЛђЛЏбЇгеБфЪжЖЮЃЌЕЭЮТЕШРызгЬхЃЈLTPЃКlow-temperature plasmaЃЉдђЪЧвЛжжаажЎгааЇЕФЮяРэ-ЛЏбЇИДКЯгеБфЗНЗЈЁЃЕШРызгЬхЪЧГ§ЙЬЬЌЁЂвКЬЌЁЂЦјЬЌЭтЕФЮяжЪЕкЫФЬЌЃЌШЫЙЄЛёЕУЗНЗЈЪЧдкСНИіЕчМЋжЎМфМгИпЕчбЙЃЌЕБДяЕНЛїДЉЕчбЙЪБЃЌЕчМЋМфЕФЦјЬхЗжзгОЭБЛЕчРыЃЌВњЩњСЫЕчзгКЭРызгЛьКЯЕФЦјЬхЃЌМДЃКЕШРызгЬхЁЃвЛАуЬѕМўЯТЃЌЗХЕчЙ§ГЬжаЫфШЛЕчзгЮТЖШКмИпЃЌЕЋжиРызгЮТЖШЕЭЃЌдђећИіЬхЯЕЮТЖШВЛИпЃЈЭЈГЃБШЛЗОГЪвЮТТдИпЃЉЃЌЯрЖдгкЬЋбєФЧбљГЪЯжЕФИпЮТЕШРызгЬхзДЬЌРДЫЕЃЌЫќвђДЫБЛГЦЮЊЕЭЮТЕШРызгЬхЁЃЕЭЮТЕШРызгЬхгыЮяжЪзїгУЗНЪНБШНЯИДдгЃЌЖјЧвзїгУВњЩњЕФЮяжЪГЩЗжвВКмЗсИЛЁЃЕШРызгЬхзїгУгкЫЎЛђКЌЫЎЮяжЪПЩвдВњЩњєЧЛљЁЂЛюадбѕздгЩЛљЕШЮяжЪОпгаНЯЧПЕФбѕЛЏадЃЌЫљвдЕБЫќзїгУгкЩњЮяЗжзгШчЕААзжЪЁЂDNAЗжзгЪБПЩв§Ц№ЗжзгЫ№ЩЫЃЌЖјЕБЫќзїгУгкЯИАћжаЕФDNAЪБЃЌШчЙћЯИАћаоИДЫ№ЩЫЕФDNAЖјГіДэЃЌдђгаПЩФмв§Ц№БфвьЁЃвђДЫLTPММЪѕПЩвдгУгкгеБфг§жжЁЃLTPгеБфЗНЗЈгааэЖргХЕуЃЌЫќВйзїМђЕЅЃЌвзгкПижЦЃЌЭЛБфТЪНЯИпЃЌВЂЧвЮоЛЏбЇгеБфКЭЗХЩфадгеБфДцдкЕФЛЗОГЮлШОКЭЧБдкЗчЯеЃЌвВВЛашвЊЬЋПегеБфЫљашвЊЕФИДдгЩшБИКЭзЪН№ЭЖШыЃЌЪЧвЛжжЬиБ№ЪЪКЯЙуЗКЪЙгУЕФгеБфЪжЖЮЁЃ
ШчНёЃЌLTPгеБфММЪѕдкЖржжЮЂЩњЮягеБфжаЖМЕУЕНЙуЗКгІгУ[8]ЁЃР§ШчЃЌЖдгкЯИОњгеБфЃЌбаОПШЫдБLTPММЪѕДІРэЕФЯИОњКИЧСЫДѓГІИЫОњЃЈEscherichia coliЃЉ[9]ЁЂАЭЪЯДзЫсИЫОњЃЈAcetobacter pasteurianusЃЉ[10]ЁЂЙШАБЫсАєзДИЫОњЃЈCorynebacterium glutamicumЃЉ[11]ЁЂВњЦјГІИЫОњЃЈEnterobacter aerogenesЃЉ[12]ЁЂвѕЙЕГІИЫОњЃЈEnterobacter cloacaeЃЉ[13]ЕШИяРМЪЯвѕадОњЃЌвВАќРЈПнВнбПцпИЫОњЃЈBacillus subtilisЃЉ[14,15]ЕШИяРМЪЯбєадОњЁЃWangЕШРћгУLTPНсКЯбЧЯѕЫсФЦДІРэДѓГІИЫОњЃЌЛёЕУСЫЗДЪН-4-єЧЛљ-L-ИЌАБЫсИпВњОњжъЃЌдк5LЗЂНЭЙожаЗЂНЭ48hЃЌИУОњжъЕФВњСПДяЕНСЫ25.4 g/L[16]ЁЃLiuЕШРћгУLTPДІРэldhAКЭpflBШБЯнаЭДѓГІИЫОњОњжъB002ЃЌдОњжъдкбсбѕЬѕМўЯТЮоЗЈРћгУЦЯЬбЬЧЃЌЕЋОгеБфДІРэЃЌLL016ЕФЭЛБфЬхЛжИДЯИАћЩњГЄКЭЦЯЬбЬЧРћгУФмСІЃЌВЂдкЗЂНЭ120аЁЪБКѓЃЌРћгУ35.0g/LЦЯЬбЬЧВњЩњСЫ25.2g/LЕФчњчъЫс[17]ЁЃWangЕШЖдзЊгаL-РЕАБЫсгеЕМЦєЖЏзгЃЈИУЦєЖЏзгПижЦКЭБэДяТЬЩЋгЋЙтЕААзЃЉЕФДѓГІИЫОњНјааLTPДІРэЃЌЭЈЙ§ЖдЭЛБфЬхПтгЋЙтМЄЛюЯИАћЕФЩИбЁКЭКѓајИДбЁЃЌЛёЕУ2жъЭЛБфИпВњОњжъЃЌЦфL-РЕАБЫсХЈЖШКЭКЌСПЗжБ№ЬсИп21.00КЭ9.05%ЃЌ18.14%КЭ10.41%[18]ЁЃLiЕШРћгУЩфЦЕДѓЦјбЙЛдЙтЗХЕчЃЈвВЪЧвЛжжLTPЗХЕчЗНЪНЃЉзїгУгкМззДУЋцпОњЃЈMethylosinus trichosporiumЃЉЃЌЛёЕУЭЛБфЬхЦфЩњГЄЫйЖШКЭЕЅМгбѕУИЛюадОњгаУїЯдЬсИп[19]ЁЃDongЕШЭЈЙ§LTPКЭЯИАћЙЬЖЈЛЏРДЬсИпМйЕЅАћОњЖдЕзЮяЕФФЭЪмадЃЌОЙ§ЩИбЁЃЌЛёЕУЕФЭЛБфОњжъmut-D3ЃЌЦфзюЪЪЕзЮяХЈЖШгЩ100 mmЬсИпЕН150 mmЃЌВњЮябЬЫсЛ§РлДяЕН189 g/LЃЌБШвАЩњаЭ(WT)ЬсИп42%[20]ЁЃдкЫПзДецОњгеБфЗНУцЃЌLTPгеБфММЪѕвВЕУЕНСЫЙуЗКгІгУЁЃР§ШчЃЌShiЕШЖдКкЧњУЙЃЈAspergillus nigerЃЉКЯГЩЦЯЬбЬЧЫсНјааLTPгеБфДІРэЃЌНсЙћЕУЕН3ИігеБфжъЃЌЦфЦЯЬбЬЧЫсВњТЪЬсИпСЫ12.1%ЁЂ15.5%КЭ32.8%[21]ЁЃZhu ЕШЭЌбљЖдКкЧњУЙНјааLTPгеБфДІРэЃЌгеБфКѓОњжъЕэЗлУИЛюадЬсИп70%ЃЌДяЕН2.2ЁС103U/ml[22]ЁЃQiangЕШРћгУLTPгеБфММЪѕДІРэШ§цпВМРУЙОњЃЈBlakeslea trisporaЃЉЃЌЪЙЦфКЯГЩЗЌЧбКьЫиЕФЫЎЦНЬсИп55%ЃЌДяЕНУППЫИЩЮяжЪ26.4mgЫЎЦН[23]ЁЃQin ЕШЖдЫПзДецОњЃЈGlarea lozoyensisЃЉНјааLTPгеБфДІРэЃЌвдЬсИпЦфКЯГЩХІФЊПЕЖЈB0ЃЈPneumocandin B0ЃЉЕФФмСІЃЌЭЈЙ§геБфЛёЕУОњжъКЯГЩСПЬсИп1.39БЖЃЌДяЕН1134mg/L[24]ЁЃLiЕШЖдИпЩНБЛцпУЙЃЈMortierella alpinaЃЉНјааLTPгеБфДІРэЃЌЦфКЯГЩЕФЛЈЩњЫФЯЉЫсЯрЖдХЈЖШДяЕН45.64%ЃЌзюИпВњСПДяЕН6.82g/L[25]ЁЃ
Ш§ЁЂКьЭтЙтЦзЩИбЁММЪѕдкгеБфг§жжжаЕФгІгУ
ЭЈЙ§ЕШРызгЬхгеБфММЪѕЕФИїжжЮяРэКЭЛЏбЇгеБфЗНЪНЃЌПЩвдЛёЕУДѓСПЕФЭЛБфОњжъЁЃЕЋЪЧЃЌШчКЮДгетаЉЭЛБфжъжаАбФПЕФадзДЕФЭЛБфжъЩИбЁГіРДЃЌетЪЧгеБфг§жжЕФЙиМќЮЪЬтжЎвЛЃЌРњРДЖМЪЧвЛЯюКФЪБКФСІЁЂФбЖШКмИпЕФЙЄзїЁЃЯждкЃЌЭЈЙ§ЛљвђзщЙЄГЬЩшМЦКЭЙЙНЈММЪѕЃЌПЩвдЛёЕУДѓСПВЛЭЌЕФЮЂЩњЮяОњжъЃЌВЂЖдЛљвђаЭНјааЬНЫї[26]ЃЌЕЋЪЧЖдетаЉЭЛБфЬхНјааБэаЭШЯЪЖМАЩИбЁвВЭЌбљЪЧвЛИіЦПОБжЦдМЮЪЬтЁЃвЛАуРДЫЕЃЌдкЩИбЁЙ§ГЬжаЃЌЖдгкШБЗІЯдЩЋЛђгЋЙтЬиадЕФЗжзгЃЌЭљЭљгУЩЋЦзЛђжЪЦзНјааЖЈСПЗжЮі[27]ЁЃЯждкЃЌЫцзХздЖЏЛЏММЪѕЫЎЦНВЛЖЯЬсИпЃЌвРРЕгквЧЦїЕФЩИбЁаЇТЪПЩвдДѓДѓЬсИпЁЃЕЋЪЧЃЌетВЛНіашвЊДѓСПзЪН№ЭЖШыЃЌзюжеЛЙЪЧашвЊбЁдёКЭНЈСЂЖдЭЛБфОњжъКЯЪЪЕФЩИбЁКЭЗжЮіЗНЗЈ[27]ЁЃЬиБ№ШєЪЧЖдЮЂЩњЮяВњЮяДњаЛЭООЖВЛЩѕСЫНтЃЌдђГЃгУЕФЩИбЁЗжЮіЗНЗЈПЩФмОЭФбвдЗЂЛгзїгУЁЃБШШчЃЌЖдгкСщжЅЖрнЦЕФЗжЮіЃЌгабаОПБэУїЃЌСщжЅЪєжаПЩФмгаЖрДя316жжЕФШ§нЦРрЛЏКЯЮяЃЌЖјетаЉЛЏКЯЮяжЎМфЮЊНсЙЙРрЫЦЮяЛђЛЅЮЊДњаЛбмЩњЮя[28]ЁЃФПЧАЮвУЧЖдетаЉДњаЛЭООЖЕФСЫНтЛЙЗЧГЃЧГЃЌНіНіЖдЩйЪ§ЛЏКЯЮяШчСщжЅЫсAЁЂСщжЅЫсTЕШЕФДњаЛЕФВПЗжЭООЖгаЫљШЯЪЖ[29]ЁЃШчКЮЖдетаЉЮяжЪНјааПьЫйЁЂзМШЗЗжЮіДгЖјЪЕЯжЭЛБфОњжъЩИбЁЃЌетОЭашвЊЗЂеЙаТЕФЩИбЁЗжЮіЗНЗЈЁЃ
ЙтбЇ-ЙтЦзбЇЗНЗЈПЩвдЖдЮЂЩњЮяЕФДњаЛВњЮяМАДњаЛЙ§ГЬНјааПьЫйЁЂЪЕЪБЁЂЮоЫ№ЙлВтгыЗжЮіЃЌвђДЫПЩФмЮЊНтОігеБфг§жжЩИбЁЮЪЬтЬсЙЉгааЇЕФМьВтгыЗжЮіЙЄОп[30]ЁЃЯждкЃЌАбЙтЦзММЪѕгІгУгкЩњЮябаОПЪЧвЛжжЧїЪЦЃЌЮвУЧПЮЬтзщЖрФъРДвВвЛжБжТСІгкЗЂеЙЩњЮяЙтЦзММЪѕ[31,32]ЁЃЩњЮяЙтЦзЪЧРћгУЩњЮяЗжзгЛђЛљЭХЕФЗЂЩфЁЂЮќЪеЛђЩЂЩфЙтЦзЬиеїРДШЗЖЈЮяжЪЕФзщГЩЁЂаджЪЁЂНсЙЙМАКЌСПЃЌВЂЖдЩњЮяаджЪМАЙ§ГЬНјааЬНВтЗжЮіЕФММЪѕ[30]ЁЃЩњЮяЙтЦзММЪѕОпгаКмЖргХЕуЃЌАќРЈЃК1ЃЉПЩЖдЩњЮябљЦЗЃЈАќРЈЩњЮяЗжзгЁЂЯИАћЁЂИіЬхЃЉНјааЪЕЪБЁЂЖЏЬЌЁЂЮоЫ№МьВтЃЛ2ЃЉПЩЪЕЯжЖрЛЏбЇзщЗжЭЌЪБПьЫйМьВтгыЗжЮіЃЛ3ЃЉВйзїМђБуЁЂбљЦЗгУСПЩйЁЂЛёЕУаХЯЂСПДѓЁЃ
дкЮЂЩњЮябаОПЗНУцЃЌКьЭтЙтЦзЃЈIRЃКinfrared spectroscopyЃЉММЪѕвбгаНЯЙуЗКЕФгІгУ[8,33]ЁЃЬиБ№ЪЧИЕРявЖБфЛЛКьЭтЙтЦзММЪѕЃЈFT-IRЃКFourier transform infrared spectroscopyЃЉЃЌПЩвдгІгУгкЮЂЩњЮяЦЗжжЕФМјЖЈКЭЗжРр[34,35]ЃЌМДРћгУКьЭтЙтЦзЕФжИЮЦЦзЧјЃЌПЩЖдВЛЭЌЕФОњжъНјааМјБ№КЭОлРрЗжЮіЁЃFT-IRММЪѕЛЙПЩгУгкЖдЮЂЩњЮяЗЂНЭЙ§ГЬЕФЖЏЬЌМьВтКЭЗжЮі[36]ЃЌЖјетвЛММЪѕгаРћгкЖдгеБфОњжжЕФЩњВњЙ§ГЬЕФЪЕЪБЖЈадЖЈСПМрПиЃЌвВПЩФмгУгкОњжжДЋДњЮШЖЈадЕФбаОПЁЃFT-IRЗНЗЈвВПЩгУгкЮЂЩњЮядкаВЦШзїгУЯТЕФЩњРэЩњЛЏМьВт[37]ЃЌетЖдбаОПЮЂЩњЮяЩИбЁжаГЃгУЕФПЙадЩИбЁЛђгЊбјШБЯнадЕФЛњРэбаОПЪЎЗжживЊЁЃГ§ДЫжЎЭтЃЌКьЭтЙтЦздкГЩЯёЗНУцЕФгІгУвВПЩгУгкЙлВтЮЂЩњЮяЕФЩњГЄКЭБфЛЏЙ§ГЬЃЌДгЖјгУгкгеБфг§жжСьгђ[32,33]ЁЃР§ШчЃЌРћгУFT-IRЮЂГЩЯёММЪѕЃЌПЩЪЕЯжЖдЮЂЩњЮяЩњГЄЗЂг§ИїЛЗНкЕФЯИАћФкзщЗжКЌСПМАЪБПеЗжВМЕФЖЏЬЌбаОПЃЌДгЖјгаРћгкЖдЭЛБфОњжъЕФДњаЛЭЛБфЮЛЕуКЭЛњжЦгшвдИќМгОЋШЗЕФХаЖЯЗжЮі[38]ЁЃMelinЕШРћгУFT-IRММЪѕЖдОІУЩфЯпЗјееДІРэЕФЮЂЧђОњНјааМьВтЃЌЙлВтЕНЗјееКѓв§Ц№ЕФЯИАћжЌжЪЙ§бѕЛЏЙ§ГЬЁЂЕААзжЪКЭЬЧРрЗЂЩњЕФаоЪЮЙ§ГЬвдМАDNAЕФЙЧМмБЛЦЦЛЕ[39]ЁЃЮвУЧПЮЬтзщгІгУКьЭтЙтЦзЯдЮЂГЩЯёММЪѕЙлВтСЫЕЅЯИАћгъЩњКьЧђдхЯКЧрЫиДњаЛЙ§ГЬ[40]ЁЃЬиБ№ЕиЃЌЮвУЧгІгУЩњЮяЙтЦзММЪѕдкгеБфОњжъбЁг§ЗНУцвВПЊеЙСЫвЛЯЕСаЕФбаОПЙЄзї[41-45]ЁЃР§ШчЃЌЮвУЧгІгУКьЭтЙтЦзКЭРТќЙтЦзЖдгъЩњКьЧђдхLTPгеБфжъНјааЗжЮі[41-44]ЁЃРћгУLTPгеБфММЪѕЃЌЮвУЧЖдгъЩњКьЧђдхНјаагеБфДІРэЃЌЛёЕУСЫДѓСПе§ЭЛБфОњжъЃЈЭЈЙ§ЕЅПЫТЁдхЯИАћЕФЗжРыКЭЩњГЄЫйТЪЕФВтЖЈЃЌНЋЩњГЄЫйТЪГЌЙ§ГіЗЂжъ20%ЖЈвхЮЊе§ЭЛБфдхжъЃЉЃЌВЂЖдетаЉЭЛБфжъЕФЯКЧрЫиКЌСПНјааВтЖЈЃЌЗЂЯжСЫЯКЧрЫиВњСПИпгкГіЗЂжъЕФM3ЁЂM14КЭM29ЕФгъЩњКьЧђдхе§ЭЛБфдхжъЃЌЦфжаЕФM3дхжъЕФгъЩњКьЧђдхЯКЧрЫиЕФВњСП3.35%w/wЃЌЮЊГіЗЂжъWTЃЈ2.11%w/wЃЉЕФ1.59БЖЁЃРћгУКьЭтЙтЦзПЩвдКмШнвзНЋе§ЭЛБфжъгыГіЗЂжъЧјЗжПЊРДЃЌВЂЧвЛљгкНќКьЭтЙтЦзЪ§ОнЮвУЧЙЙНЈСЫЯКЧрЫиКЌСПЗжЮіЕФЖЈСПФЃаЭ[41-43]ЃЌЦфзМШЗПЩППадЩѕжСПЩгыГЃЙцЖЈСПЗНЗЈЯрБШНЯЁЃвдЯТЗжБ№НщЩмЮвУЧдкЕЭЮТЕШРызгЬхгеБфСщжЅММЪѕКЭЙтЦзЩИбЁЗжЮіЗНЗЈЩЯЫљзіЕФвЛаЉбаОПЙЄзїЁЃ
ЫФЁЂЕШРызгЬхгеБфСщжЅММЪѕКЭЙтЦзЩИбЁЗжЮіЗНЗЈ
1ЁЂРћгУЕШРызгЬхгеБфСщжЅдЩњжЪЬхЛёЕУЖрЬЧКЌСПИпЕФгеБфОњжъ[46]
ЮЊСЫЪЕЯжгааЇЕФLTPгеБфЃЌЮвУЧздМКДюНЈСЫНщжЪзшЕВЕШРызгЬхЗХЕчЃЈDBDЃЉЗЂЩњзАжУЃЌВЂРћгУетИізАжУЖдСщжЅдЩњжЪЬхНјаагеБфЁЃЮвУЧЪЕбщгУЕФСщжЅЪЧГрСщжЅЃЈGanoderma lucidumЃЉЁЃЪЕбщзАжУШчЭМ1ЫљЪОЃЌЮвУЧЩшМЦЕФLTPгеБфЩшБИВњЩњЕШРызгЬхЪЧЭЈЙ§НщжЪзшЕВЗХЕчЃЈDBDЃКdielectric barrier dischargeЃЉЗНЪНЃЌЗХЕчЕчдДВЩгУФЩУыТіГхИпбЙЕчдДЃЌЗХЕчЦјЬхПЩвдВЩгУыВЦјЁЂЕЊЦјЕШЃЌгЩЦјБэПижЦСїСПзЂШызджЦЕФдВжљаЮШнЦїЃЌФкжУЪЏгЂЗДгІУѓЃЌзЂШыСщжЅдЩњжЪЬхШмвКЃЌЩЯЗНЙЬЖЈПЩЕїНкИпЖШЕФЕчМЋЃЌЕчМЋСЌНгИпбЙФЩУыТіГхжБСїЕчЁЃLTPЗХЕчаЇЙћПЩЭЈЙ§ЦјЬхжжРрЁЂЦјЬхСїСПКЭЕчбЙЧПЖШЕШЕїПиЁЃдкСщжЅгеБфЪЕбщжаЃЌЮвУЧашвЊгХЛЏLTPЗХЕчЪЕбщЬѕМўЁЃЪЕбщжаВЩгУЕФЙЄзїЦјЬхЃКHeЦјЃЌЗХЕчЕчбЙЃК13kVЃЌЦјСїСПЃК2L/minЃЌЗХЕч4minЁЃ

ЭМ1НщжЪзшЕВЃЈDBDЃЉЕЭЮТЕШРызгЬхЃЈLTPЃЉЗЂЩњзАжУЪОвтЭМ
РћгУLTPгеБфСщжЅдЩњжЪЬхжЎКѓЃЌЮвУЧгУRAPD(random amplified polymorphic DNAЃЉМДЫцЛњРЉдіЖрЬЌадDNAБъМЧЖдДІРэЙ§ЕФдЩњжЪЬхНјааГѕВНЗжЮіЃЌМДгУЫќПЩвдМьВтЪЧЗёгаDNAЕФИФБфДгЖјХаЖЯЪЧЗёгеБфВњЩњ[41]ЁЃЭЈЙ§ЖрДЮRAPDМьВтЃЌЮвУЧХХГ§СЫМйвѕадКЭМйбєадЧщПіЃЌДгЖјШЗЖЈСЫLTPгеБфОњжъЁЃЭЌЪБЃЌЮвУЧЭЈЙ§СђЫсньЭЊЗНЗЈ[47]ВтСПСщжЅЖрЬЧКЌСПЃЌПМВьгеБфжъЖрЬЧКЌСПЕФИФБфЁЃНсЙћШчЭМ2ЫљЪОЁЃ
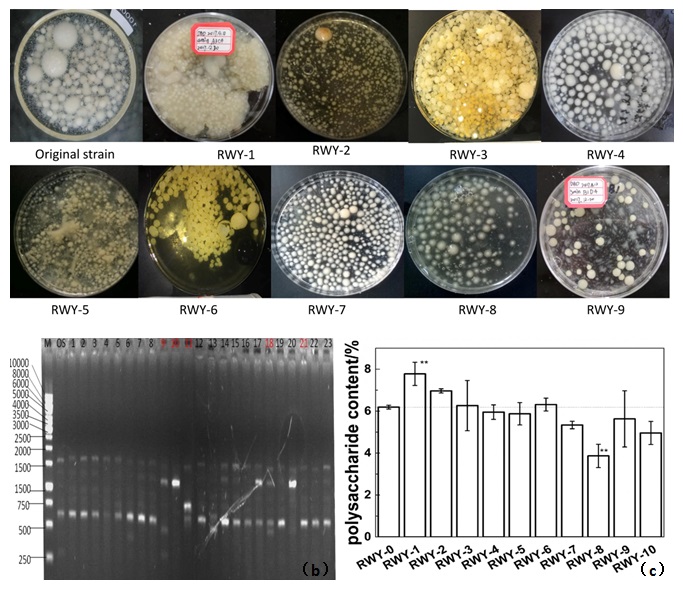
ЭМ2 геБфСщжЅОњжъзДЬЌЁЃЃЈaЃЉВЛЭЌгеБфСщжЅОњЫПЧђЕФБэЙлаЮЬЌЃЛЃЈbЃЉВПЗжLTPгеБфОњжъRAPDЕчгОЭМЃЛЃЈcЃЉВПЗжгеБфОњжъЖрЬЧКЌСПБШНЯ
гЩЭМ2ПЩвдПДГіЃЌгеБфСщжЅОњжъОњЫПЧђДѓаЁЁЂаЮЬЌЁЂбеЩЋЁЂЩњГЄЫйЖШЁЂЩњЮяСПЕШДцдкзХБШНЯУїЯдЕФВюБ№ЁЃРћгУЖрИів§ЮязщНјааRAPDЕчгОЗжЮіЃЌВПЗжгеБфОњжъЕФЕчгОЬѕДјгыOSГіЗЂжъГіЯжСЫЯджјЕФВювьЃЌШЗЖЈЪЧЗЂЩњЭЛБфЕФОњжъЁЃетаЉЭЛБфжъЯрБШгкГіЗЂжъЃЌЦфСщжЅЖрЬЧКЌСПвВгаСЫКмДѓВювьЁЃRWY-1ЁЂRWY-2ЁЂRWY-3КЭRWY-6ЦНОљЖрЬЧКЌСПИпгкГіЗЂжъЃЛRWY-4ЁЂRWY-5ЁЂRWY-7ЁЂRWY-8ЁЂRWY-9КЭRWY-10ЦНОљЖрЬЧКЌСПЕЭгкГіЗЂжъЁЃRWY-1КЭRWY-8ЖрЬЧКЌСПгыГіЗЂжъЯрБШгаЯджјВювьЁЃПЩвдПДЕНЃЌОЙ§LTPгеБфММЪѕДІРэЃЌВПЗжОњжъЕФЖрЬЧКЌСПгыГіЗЂжъЯрБШгаСЫЯджјИФБфЁЃ
2ЁЂРћгУКьЭтЙтЦзЗНЗЈМјБ№ЖрЬЧКЌСПИпЕФгеБфОњжъ[48]
ЮвУЧЭЈЙ§LTPЕУЕНДѓСПСщжЅгеБфЭЛБфжъЁЃЮЊСЫЖдетаЉгеБфжъНјааЩИбЁЗжЮіЃЌвЛЗНУцЮвУЧдкбаОПжаМЬајВЩгУДЋЭГЕФВтСПЗНЗЈЃЌЖдетаЉОњжъЕФСщжЅЖрЬЧКЭШ§нЦКЌСПНјааВтСПКЭЗжЮіЃЌСэвЛЗНУцЃЌЮвУЧвВдкНЈСЂКЭЗЂеЙЛљгкЙтЦзЕФЗжЮіЗНЗЈЃЌвдБугкЗЂеЙаТЕФгеБфг§жжЩИбЁЙЄОпгыЪжЖЮЁЃетРяжївЊНщЩмЮвУЧЖдСщжЅОњЫПЬхКьЭтЙтЦзНјаабаОПЕФЙЄзїЁЃ
КьЭтЙтЦзПЩвдБэеїЗжзгЬиЖЈЙІФмЛљЭХвдМАећИіЗжзгНсЙЙЕФИїжжЬиеїЁЃЩњЮяЬхзїЮЊИДдгЕФЯЕЭГЃЌжївЊгЩЫЎЁЂжЌЗОЁЂЕААзжЪЁЂЬМЫЎЛЏКЯЮяКЭЦфЫћГЩЗжзщГЩЁЃдкЩњЮяЛњЬхжаетаЉзщЗжЕФКЌСПЁЂзщГЩЁЂЗжзгНсЙЙМАЗжзгФкКЭЗжзгМфЕФЯрЛЅзїгУЃЌИГгшСЫЩњЮяЬхгаБ№гкЦфЫќЮяЬхЕФЙтЦзбЇаджЪЁЃвђДЫжаКьЭтЙтЦзМШПЩвдЬсЙЉНсЙЙ-ЙІФмЙиЯЕЕФаХЯЂЃЌгжПЩвдзїЮЊЛЏбЇГЩЗжЖЈСПЗжЮіЕФЙЄОп[34]ЁЃНќКьЭтЙтЦзЪЧгЩКьЭтжаЕФЛљБОеёЖЏдОЧЈЕФБЖЦЕКЭзщКЯЦЕЙЙГЩЕФЃЌЦфЮќЪеЯЕЪ§вЛАуБШжаКьЭтЙтЦзЕЭ10~10000БЖЁЃвђДЫЃЌгыжаКьЭтЙтЦзВЛЭЌЃЌНќКьЭтЙтЦзашвЊВЩМЏДѓСПЙтЦзЃЌжївЊгУгкЩњЮябљБОЕФЖЈСПЗжЮіЃЌЖјЦфжавЛИіЙиМќВНжшЪЧНЈСЂвЛИіаЃе§МЏФЃаЭЃЌГЃгУЕФЛЏбЇМЦСПбЇЗНЗЈАќРЈЖрдЊаЃе§ММЪѕЃЌШчзюаЁЖўГЫЗЈЃЈPLSЃЉЁЂЖрдЊЯпадЛиЙщЃЈMLRЃЉЁЂжїГЩЗжЛиЙщЃЈPCRЃЉЁЂОжВПМгШЈЛиЙщЃЈLWRЃЉЕШ[42,44]ЁЃ
дкКьЭтЙтЦзВтСПЗжЮіЕФЪЕбщжаЃЌЮвУЧНЋЗЂНЭКѓЕФГіЗЂжъКЭгеБфжъЕФСщжЅОњЫПЬхЧхЯДКѓРфЖГИЩдяЃЌбаГЩЗлФЉзДЙ§ЩИБИВтЁЃИљОнКьЭтЙтЦзВЈЖЮПЩвдЗжЮЊжаКьЭтЃЈmid-IRЃЌЦзЧјЗЖЮЇЮЊ4000-200 cm-1ЃЉКЭНќКьЭтЃЈnear-IRЛђNIRЃЌ ЦзЧјЗЖЮЇЮЊ12500ЁЋ4000cm-1ЃЉВтСПЁЃдкжаКьЭтЙтЦзМьВтжаЃЌЮвУЧАббљЦЗбЙЦЌжУгкBruker Alpha-TКьЭтЙтЦзвЧжаЙтЦзЩЈУшЁЃЩЈУшЗЖЮЇЃК4000-400cm-1ЃЌЗжБцТЪ4cm-1ЃЌЩЈУш64ДЮЃЌOPUS 7.0Ъ§ОнДІРэШэМўЪеМЏНсЙћВЂЗжЮіЁЃдкНќКьЭтЙтЦзМьВтжаЃЌЮвУЧАбСщжЅОњЫПЬхжЦГЩЗлФЉЃЌгУЕТЙњВМТГПЫЙЋЫОMPAаЭНќКьЭтЙтЦзвЧВЩМЏЦфНќКьЭтТўЗДЩфЙтЦзЃЌВЈЪ§ЗЖЮЇЮЊ12500-400cm-1ЃЌЩшжУЙтЦзЗжБцТЪЮЊ16cm-1ЃЌЩЈУш32ДЮЁЃЦНааМьВт2ДЮШЁЦНОљЙтЦзЗжЮіЁЃМЧТМЕФЕфаЭЙтЦзШчЭМ4ЫљЪОЁЃ
ЭМ4 СщжЅОњЫПЬхЕФКьЭтЙтЦзЃЈАќРЈжаКьЭтКЭНќКьЭтЙтЦзЃЉМьВтСїГЬЪОвтЭМ
ЃЈ1ЃЉСщжЅОњЫПЬхЕФжаКьЭтЃЈmid-IRЃЉЬиеїадЙтЦз
ЕфаЭЕФСщжЅИЩдяОњЫПЬхКЭгеБфСщжЅОњЫПЬхЕФFT-IRЙтЦзВтСПНсЙћШчЭМ6aЫљЪОЃЌЦфЕфаЭЗхЮЛЮЛгк1425 cm-1 [49,50]ЁЂ1316 cm-1 [51]ЁЂ1152 cm-1 [52]ЁЂ1078 cm-1 [53]ЁЂ1025 cm-1 [54,55]КЭ951 cm-1[55,56]ЕШЮЛжУОљгаБШНЯЧхЮњЕФЮќЪеЗхЃЌЙщЪєЧщПіМћБэ1ЁЃ
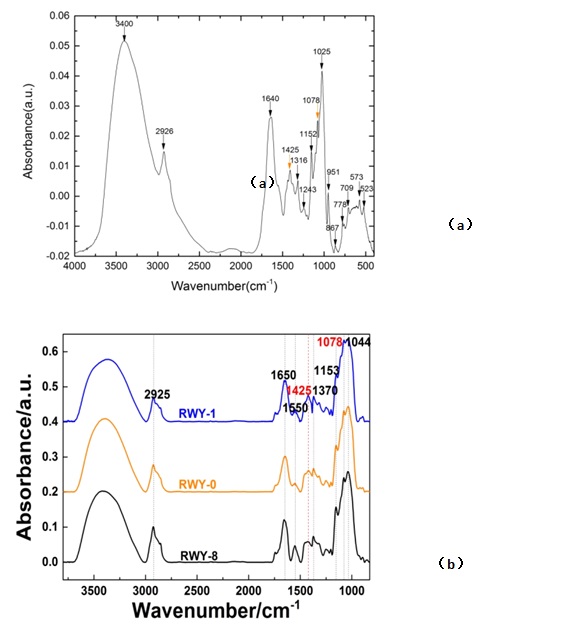
ЭМ6 ЃЈaЃЉЕфаЭЕФСщжЅОњЫПЬхЕФКьЭтЙтЦзЃЛЃЈbЃЉВЛЭЌгеБфСщжЅОњжъКьЭтЙтЦзБШНЯЃЈЖрЬЧКЌСПВЛЭЌЃЉ
Бэ 1 СщжЅИЩдяОњЫПЬхжаКьЭтЬиеїЙтЦзЙщЪє
ВЈЪ§(cm-1) | ЛљЭХЙщЪє |
3400 | -OH ЩьЫѕеёЖЏ[57] |
2926 | CH2ЗЧЖдГЦЩьЫѕеёЖЏ[58] |
1640 | ѕЃАЗ I[58] |
1457 | ЖрЬЧжаЕФ-CH2[59] |
1425 | ФОжЪЫиКЭЬМЫЎЛЏКЯЮяжаЕФC-HБфаЮеёЖЏ[49,60] |
1372 | УцФкЭфЧњеёЖЏC-H [61,62] |
1314-1316 | ЯЫЮЌЫижа-CH2ЭфЧњеёЖЏ[51] |
1243 | УцФк-COH/CHЭфЧњеёЖЏ[61] |
1152-1156 | ЬЧмеМќC-O-CЗЧЖдГЦЩьЫѕеёЖЏ[52] |
1078 | ІТ-ЬЧмеМќC-OЩьЫѕеёЖЏ[50,53,54] |
1044 | C-O-CЛљЭХЩьЫѕеёЖЏ[63] |
1025 | ІС-ЬЧмеМќC-OЩьЫѕеёЖЏ[54,55] |
951 | ІТ-ЬЧмеМќ [55]; C-O КЭ C-C ЩьЫѕеёЖЏ[56] |
867 | ІУ (C-H)[64]; пЛрЋЛЗ[65] |
778 | COO- БфаЮеёЖЏ[66] |
709 | CHУцЭтЭфЧњ[67] |
573 | ЖрЬЧЛЗЭфЧњеёЖЏ[68] |
523 | пСрЋЛЗ[65] ; C=OЗЧЖдГЦБфаЮеёЖЏ[69] |
ЮЊСЫМјБ№жаКьЭтЙтЦзжагыСщжЅЖрЬЧЕФЬиеїЙтЦзЃЌЮвУЧЛЙЖдКЌВЛЭЌСщжЅЖрЬЧЕФСщжЅбљЦЗНјааЙтЦзБШНЯКЭЗжЮіЁЃДгжаКьЭтЙтЦзжаЃЌЮвУЧПЩвдЪЖБ№ГіЬМЫЎЛЏКЯЮяЕФЬиеїДјЃЈЭМ6aЃЉЃЌМДдк1425 cm-1КЭ1078 cm-1ДІЦзЗхЃЌЫќУЧдкЖрЬЧКЌСПНЯИпЕФбљЦЗжаЯрЖдЧПЖШНЯЧПЃЈЭМ6bЃЉЁЃ
ЃЈ2ЃЉНќКьЭтЙтЦзЃЈNIRЃЉВтЖЈСщжЅОњЫПЬхжаЖрЬЧЕФКЌСП
ЮЊСЫНјааСщжЅЖрЬЧЖЈСПЗжЮіЃЌЮвУЧЛЙЪеМЏДѓСПВЛЭЌСщжЅОњжъЖджЎНјааНќКьЭтЙтЦзВтСПЁЃСщжЅИЩдяОњЫПЬхЕФЕфаЭНќКьЭтЙтЦзШчЭМ7aЫљЪОЃЌЦфжа8403 cm-1ЃЌ6896 cm-1ЃЌ5155 cm-1ДІЕФЮќЪеЗхЙщЪєЮЊЫЎЃЌЖјдк4307 cm-1ЃЌ4405 cm-1ЃЌ5787 cm-1КЭ5935 cm-1ДІЕФЮќЪеЗхЪєгкЬМЫЎЛЏКЯЮяЮќЪеЗхЁЃНќКьЭтЙтЦзЬиеїЦзЕФЯъЯИЙщЪєВЮМћБэ2ЁЃЮвУЧРћгУНќКьЭтЙтЦзЪ§ОнПЩвдЙЙНЈЦфЖрЬЧКЌСПЕФЖЈСПФЃаЭЃЌЭЈЙ§БШНЯЃЌЮвУЧЗЂЯжЕБбЁШЁЕФЙтЦзЗЖЮЇШчЭМ8ЃЈaЃЉКЭЃЈbЃЉЫљЪОЃЌдЄДІРэЗНЗЈЮЊвЛНзЕМЪ§ЃЌЕУЕНСщжЅОњЫПЖрЬЧЖЈСПФЃаЭНЯКУЁЃзїЮЊаЃе§МЏЃЌдкЮЌЪ§=6ЪБЃЌЦфЖЈСПФЃаЭЕФОіЖЈЯЕЪ§R2=0.9779ЃЌНЛВцбщжЄОљЗНИљЃЈRMSECVЃЉ=0.467ЃЌВаСєдЄВтЦЋВюжЕЃЈRPDЃЉ=6.73ЃЌНсЙћШчЭМ8ЃЈcЃЉЫљЪОЁЃЮвУЧРћгУИУЖЈСПФЃаЭЖдLTPгеБфЛёЕУЕФСщжЅОњжъНјааЖЈСПЗжЮіЃЌНсЙћШчЦфRMSEP=0.445ЃЌЯрЙиЯЕЪ§=0.9684ЃЌНсЙћШчЭМ8ЃЈdЃЉЫљЪОЁЃ

ЭМ 7ЃЈaЃЉ СщжЅИЩдяОњЫПЬхНќКьЭтЙтЦзЖЈСПФЃаЭбЁдёЧјЖЮЃЛЃЈbЃЉбЁдёЧјЖЮвЛНзЕМЪ§ЃЛЃЈcЃЉ76ИіОњжъЙЙНЈЕФаЃе§МЏЃЛЃЈdЃЉDBDгеБфОњжъдЄВтМЏ
Бэ 2 СщжЅОњЫПЬхНќКьЭтЙтЦзЙщЪє
ВЈЪ§ (cm-1) | ЙтЦзЙщЪє | жївЊРДдД(s) |
8403 | O-HЩьЫѕеёЖЏЕФвЛМЖБЖЦЕКЭO-HЭфЧњеёЖЏЕФзщКЯЦЕ (2ІЭ1,3+ІЭ2) | ЫЎ[70] |
6896 | ЫЎ [70] | |
6674 | O-HЩьЫѕеёЖЏЕФвЛМЖБЖЦЕ | |
6307 | O-HЩьЫѕеёЖЏЕФвЛМЖБЖЦЕ | ДМЛђЫЎ[70] |
5935 | CЈCHЩьЫѕеёЖЏЕФвЛМЖБЖЦЕ | ФОжЪЫи[71], АыЯЫЮЌЫи [71, 72] |
5787 | -CH2жаCЈCHЕФвЛМЖБЖЦЕ | ЬМЫЎЛЏКЯЮя [73] |
5155 | ЫЎжаO-HЩьЫѕеёЖЏКЭБфаЮеёЖЏЕФзщКЯЦЕ | ЫЎ [74] |
4719 | O-H КЭ C-O ЩьЫѕеёЖЏЕФзщКЯЦЕ | ЖрЬЧ [75] |
4405 | O-H ЩьЫѕеёЖЏКЭ C-OЩьЫѕеёЖЏЕФзщКЯЦЕ | ЖрЬЧ [76] |
4307 | C-H ЩьЫѕеёЖЏКЭC-H2БфаЮеёЖЏЕФзщКЯЦЕ | ЖрЬЧ [70] |
4021 | C-HЩьЫѕеёЖЏКЭ C-CЩьЫѕеёЖЏЕФзщКЯЦЕ | ЯЫЮЌЫи [70] |
етРяВЙГфЫЕУївЛЯТЙигкСщжЅЖрЬЧНќКьЭтЙтЦзЗжЮіКЭЖЈСПФЃаЭЕФКЯРэадКЭПЩППадЁЃЖдгкСщжЅЖрЬЧКЌСПИпЕЭВЛЭЌОњжъдк4307 cm-1КЭ4405 cm-1ДІНќКьЭтЙтЦзЗхЮЛгаБШНЯЯджјЕФВювьЃЌШчЭМ8ЃЈaЃЉЫљЪОЁЃЖјЮвУЧЕФСПЛЏФЃаЭКИЧСЫ5268.8-4000 cm-1ЕФЙтЦзЗЖЮЇЃЌЦфжаАќРЈдк4307КЭ4405 cm-1етСНДІЬиеїадЗхЮЛЃЌетСНИіЗхЮЛЖдгІзХжаКьЭтЕФ1425КЭ1078 cm-1ЗхЮЛЁЃЖј1425КЭ1078 cm-1ЗхЮЛЗжБ№ЖдгІгкЦЯОлЬЧпСрЋЛЗЕФC-HЭфЧњКЭC-O-HЭфЧњеёЖЏЃЌЦЯОлЬЧЙуЗКДцдкгкВЛЭЌжжРрЕФецОњжа[77]ЁЃОпЬхЖјбдЃЌ1078 cm-1ЪЧФОжЪЫиКЭЬМЫЎЛЏКЯЮяЕФІТ-ЦЯОлЬЧЕФC-OЩьЫѕеёЖЏ[50]ЃЌетгыНќКьЭтЙтЦзжаЕФ4405 cm-1ЯрЖдгІЃЌвђЮЊ4405 cm-1ЪЧгЩO-HЕФЩьЫѕеёЖЏКЭC-OЩьЫѕеёЖЏЕФзщКЯЦЕЁЃ1425 cm-1ЗхЮЛЖдгІгкФОжЪЫиКЭЬМЫЎЛЏКЯЮяжаЕФC-HЕФБфаЮеёЖЏ[49]ЃЌетгыНќКьЭтЙтЦзжаЕФ4307 cm-1ЗхгаЙиЃЌетЪЧгЩгкC-HЩьЫѕеёЖЏКЭCH2БфаЮеёЖЏзщКЯЖјГЩЁЃЮЊСЫНјвЛВНбщжЄетвЛЕуЃЌЮвУЧЛЙМЦЫуСЫжаКьЭтЙтЦзКЭНќКьЭтЙтЦзЕФЖўЮЌЯрЙиЙтЦзЃЌгЩДЫбщжЄСщжЅОњЫПЬхЖрЬЧНќКьЭтЙтЦзКЭжаКьЭтЙтЦзжЎМфЕФЙиЯЕЃЌНсЙћШчЭМ8ЃЈbЃЉЫљЪОЁЃгЩЭМжаПДЕНЃЌНќКьЭтЙтЦзЗЖЮЇ(5268.8~4000 cm-1)гыжаКьЭтЙтЦз(1422~1376 cm-1)ЕФЯрЙиадзюЧПЃЌете§ЪЧРДздСщжЅЖрЬЧГЩЗжЕФЙБЯзЁЃ
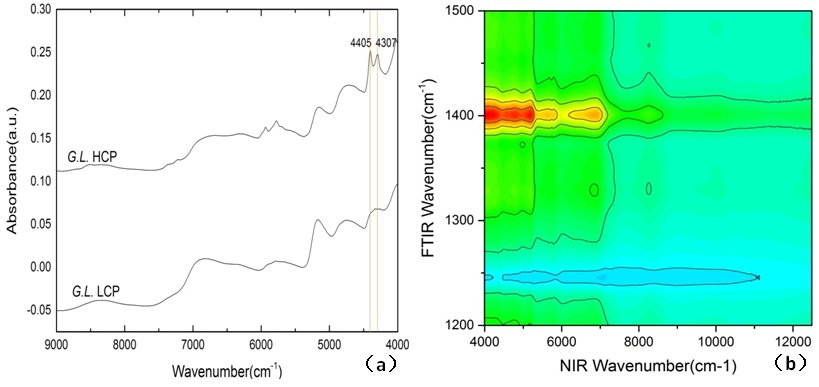
ЭМ8 ЃЈaЃЉСщжЅЖрЬЧКЌСПИпЃЈG.L. HCPЃЉЕЭЃЈG.L.LCPЃЉОњжъНќКьЭтЙтЦзЃЛЃЈbЃЉСщжЅОњжъжаКьЭтКЭНќКьЭт2DЯрЙиЙтЦз
ЃЈ3ЃЉКьЭтЙтЦзМАРТќЙтЦзММЪѕдкСщжЅШ§нЦЗжЮіжаЕФгІгУ
СэвЛЗНУцЃЌЖдгкСщжЅШ§нЦГЩЗжМАКЌСПЕФВтСПгыЗжЮіЃЌЮвУЧвВВЩгУЙтЦзЗжЮіЗНЗЈзіСЫвЛаЉЬНЫїадЕФЙЄзї[78,79]ЁЃСщжЅШ§нЦжжРрЖрЖјИДдгЃЌМјБ№Ц№РДЯрЖдБШНЯРЇФбЁЃвдЭљбаОПжаЃЌЙигкСщжЅШ§нЦЯдЩЋЗжЮіЗНЗЈПЩФмДцдквЛаЉЮЪЬтЃЌЖдДЫЮвУЧзіЙ§вЛаЉЬНЬж[80]ЁЃ
ЖдгкЙтЦзбаОПЙЄзїЃЌЮвУЧашвЊЕУЕНвзгкЙтЦзЗжЮіИїжжСщжЅШ§нЦБъзМбљЦЗЁЃвЛАуШ§нЦПЩДгСщжЅзгЪЕЬхжаЬсШЁЃЛЕЋдквКЬхЗЂНЭЬѕМўЯТЃЌСщжЅОњЫПвВЛсВњЩњвЛЖЈСПЕФСщжЅШ§нЦЃЌжЛЪЧВЛЭЌОњжъОњЫПЬхСщжЅШ§нЦКЌСПгаЫљВЛЭЌЁЃвЛАуРДЫЕЃЌРћгУHPLCЗжЮіЃЌЮвУЧПЩвдСщжЅШ§нЦНјааГѕВНЗжЮіЃЌПЩвдПДЕНЃЌВЛЭЌОњжъЕФСщжЅШ§нЦКЌСПВювьКмДѓЃЌНсЙћШчЭМ9ЫљЪОЃЌЦфжаШчG058КЭG009ЯджјИпгкG080ЁЃ
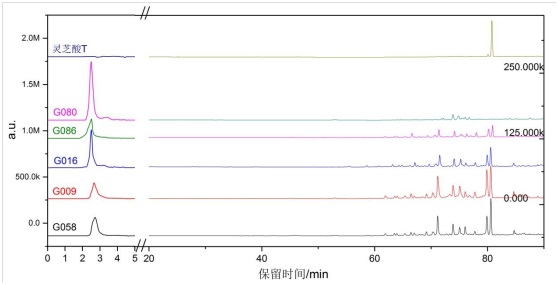
ЭМ9 HPLCВтСПВЛЭЌСщжЅОњЫПЬхжаШ§нЦКЌСП
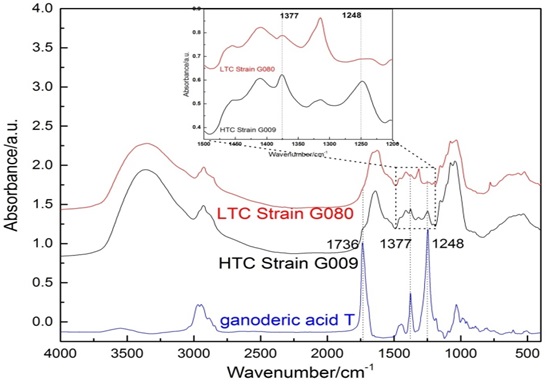
ЭМ10 ВЛЭЌШ§нЦКЌСПЕФСщжЅОњжъЕФКьЭтЙтЦзБШНЯ
НјвЛВНПЩвдгУИїжжЬсДПжЦБИИїжжШ§нЦДПЦЗЃЌвдБугкбАевШ§нЦЙтЦзЬиеїЗхЁЃЮвУЧБШНЯСЫG009гыG080ЕФжаКьЭтЙтЦзЃЌШчЭМ10ЫљЪОЁЃНЋСщжЅОњЫПЬхжаШ§нЦКЌСПИпЕЭВЛЭЌЕФОњжъгыСщжЅЫсTЕФКьЭтЙтЦзЖдБШЃЌЮвУЧЗЂЯжЦфжаЕФЬиеїадЗхЮЛЗжБ№ЮЛгк1736ЁЂ1377КЭ1248cm-1ЁЃЦфжа1736 cm-1ЖдгІзХC=OЩьЫѕеёЖЏЃЌ1377 cm-1ЖдгІгкCH3жаC-HЖдГЦЭфЧњеёЖЏЃЌ1248 cm-1ЖдгІгкC-HУцФкеёЖЏЁЃДгЭМжаПЩвдПДГіЃЌШ§нЦКЌСПИпЕФОњжъG009дкЩЯЪіЗхЮЛБШG080ИќЮЊУїЯдЁЃетаЉЙтЦзЗхЮЛЖдСщжЅОњЫПЬхжаШ§нЦКЌСПЖЈадЗжЮіжагаживЊзїгУЃЌЕЋвЊОпЬхЗжЮіетаЉЙтЦзЃЌЛЙашвЊЖдЙтЦзНјааЯъОЁЕФРэТлМЦЫуЁЃзюНќЃЌЮвУЧЖдВЛЭЌСщжЅЫсЕФжаКьЭтКЭРТќЙтЦзЗхЮЛНјааСЫDFTМЦЫуЃЌЯъЯИЗжЮіМЦЫуСЫВЛЭЌжжРрСщжЅЫсЕФЬиеїадЗхЮЛКЭЙтЦзЬиеї[79]ЁЃЕЋЪЧвђЮЊШ§нЦжжРрКмЖрЃЌИїжжСщжЅОњжъШ§нЦжжРрМАКЌСПвВДцдкКмДѓВювьЃЌЫљвдЙигкСщжЅШ§нЦИќЮЊЯЕЭГЕФЙтЦзбаОПЙЄзїЛЙдкНјаажЎжаЁЃ
ЮхЁЂеЙЭћ
СщжЅзїЮЊДЋЭГЕФвЉгУОњжаВнвЉВФЃЌОпгаЦфСщжЅЖрЬЧКЭШ§нЦЪЧвбжЊЕФвЉаЇГЩЗжЁЃЮЊСЫЬсИпСщжЅОњжъЕФСщжЅвЉгУГЩЗжЃЌПЩвдВЩгУЕЭЮТЕШРызгЬхЕШгеБфЗНЪНЖддЪМГіЗЂжъНјаагеБфДІРэЃЌДгЖјЛёЕУДѓСПЭЛБфЬхПтЃЌВЂДгжаЩИбЁСщжЅЖрЬЧЁЂШ§нЦЛђЦфЫќгааЇГЩЗжКЌСПИпЕФОњжъЃЌгЩДЫЗсИЛСщжЅЕФжжжЪзЪдДЃЌДяЕНгХСМЦЗжжбЁг§ЕФФПЕФЁЃЮвУЧбаОПБэУїЃЌАбЕЭЮТЕШРызгЬхММЪѕКЭЙтЦзММЪѕгаЛњНсКЯЦ№РДЃЌетЪЧвЛжжаажЎгааЇЕФЭООЖЁЃФПЧАЃЌдкЕЭЮТЕШРызгЬхММЪѕбаОПЗНУцЃЌГ§СЫгХЛЏгеБфВЮЪ§вдЭтЃЌЛЙашвЊЖдецОњгеБфЛњРэНјвЛВНбаОПЁЃдкЙтЦзбаОПЗНУцЃЌвЛЗНУцвЊгыДЋЭГЗжЮіЗНЗЈЛЅЮЊбщжЄЃЌСэвЛЗНУцвЊЭЈЙ§РэТлМЦЫуНјааЙтЦзНтЮіЁЃдкКьЭтЙтЦзгІгУЗНУцЃЌПЩвдАбжаКьЭтКЭНќКьЭтЙтЦзММЪѕНсКЯЦ№РДЃЌжївЊРћгУжаКьЭтПЩвдЖЈадЗжЮіЃЌРћгУНќКьЭтНјааЖЈСПНЈФЃЃЌетЖдгкЗЂеЙЦЗжжжЪСППьЫйМјБ№КЭЩИбЁЁЂНЈСЂХкЙцБъзМПЩФмЗЧГЃгагУЃЌОпгаКмКУЕФгІгУКЭЗЂеЙЧАОАЁЃДЫЭтЃЌКьЭтЙтЦзЛЙПЩвдНсКЯРТќЙтЦзНЈСЂСщжЅвЉгУГЩЗжЕФЖЈадКЭЖЈСПЗжЮіФЃаЭЁЃР§ШчЃЌдкШ§нЦЗжЮіЗНУцЃЌЮвУЧПЩвдРћгУРТќЙтЦзЗжЮіКЭМјБ№ВЛЭЌШ§нЦжжРрЃЌНјвЛВННЈСЂЛљгкЙтЦзММЪѕЕФВњЦЗЗжЮіКЭМьбщБъзМЁЃзмжЎЃЌЮвУЧетЯюбаОПВЛНігаРћгкСщжЅг§жжЃЌЖјЧвЛЙПЩвдЭЦЙуЕНЦфЫќЪГгУОњКЭвЉгУОњЃЌР§ШчЖдзЯжЅЁЂХЃеСжЅЁЂЩЃЛЦЕШНќдДвЉгУЫПзДецОњНјааЕШРызгЬхгеБфМАЙтЦзЩИбЁЃЌвдЦкЛёЕУИќКУЕФЦЗжжЃЌвдЭЦЖЏЮвЙњЪГвЉгУецОњг§жжвЕЕФЗЂеЙЁЃ
ВЮПМЮФЯз
[1] PENG R, FU Y Z, ZOU J, et al. Improvement of polysaccharide and triterpenoid production of Ganoderma lucidum through mutagenesis of protoplasts [J]. Biotechnol Biotechnol Equip, 2016, 30(2): 381-387.
[2] CHOONG Y K, CHEN X D, JAMAL J A, et al. Preliminary Results of Determination of Chemical Changes on Lingzhi or Reishi Medicinal Mushroom, Ganoderma lucidum (W.Curt.:Fr.)P. Karst. (Higher Basidiomycetes) Carried by Shenzhou I Spaceship with FTIR and 2D-IR Correlation Spectroscopy [J]. International Journal of Medicinal Mushrooms, 2012, 14(3): 295-305.
[3] ЕЫДККЃ, ЛЦЭЂюк, МНБІйјЃЌЕШ. ЬЋПегеБфЖдСщжЅОњжъЬиадЕФгАЯь [J]. ЪГгУОњ, 1999, 05): 9-10.
[4] еХвјЦМ, еХЪц, ЭєбѓЃЌЕШ. СщжЅдЩњжЪЬхЕФИпаЇжЦБИМАгеБф [J]. АВЛеХЉвЕПЦбЇ, 2012, 40(25): 12379-12381.
[5] Жгёчт, УчОДжЅ, ВмдѓКчЃЌЕШ. СщжЅдЩњжЪЬхЛЏбЇгеБфг§жж [J]. ЪГЦЗбаОПгыПЊЗЂ, 2012, 33(06): 166-170.
[6] WEI Z H, DUAN Y Y, QIAN Y Q, et al. Screening of Ganoderma strains with high polysaccharides and ganoderic acid contents and optimization of the fermentation medium by statistical methods [J]. Bioprocess and Biosystems Engineering, 2014, 37(9): 1789-1797.
[7] HAO Q, HAN X, ZOU G, et al. CRISPR-Cas9 assisted gene disruption in the higher fungus Ganoderma species [J]. 2017, 56(57-61.
[8] OTTENHEIM C, NAWRATH M, WU J C. Microbial mutagenesis by atmospheric and room-temperature plasma (ARTP): the latest development [J]. Bioresources and Bioprocessing, 2018, 5(1): 1-14.
[9] MA J F, WU M K, ZHANG C Q, et al. Coupled ARTP and ALE strategy to improve anaerobic cell growth and succinic acid production by Escherichia coli [J]. Journal of Chemical Technology & Biotechnology, 2016, 91(3): 711-717.
[10] WU X, WEI Y, XU Z, et al. Evaluation of an Ethanol-Tolerant Acetobacter pasteurianus Mutant Generated by a New Atmospheric and Room Temperature Plasma (ARTP) [J]. Journal of Bacteriology, 2015, 187(13): 4671.
[11] CHENG G, XU J, XIA X, et al. Breeding L-arginine-producing strains by a novel mutagenesis method: Atmospheric and room temperature plasma (ARTP) [J]. Prep Biochem Biotechnol, 2015, 46(5): 509-516.
[12] LU Y, WANG L, MA K, et al. Characteristics of hydrogen production of an Enterobacter aerogenes mutant generated by a new atmospheric and room temperature plasma (ARTP) [J]. Biochemical Engineering Journal, 2011, 55(1): 17-22.
[13] HUA X, WANG J, WU Z, et al. A salt tolerant Enterobacter cloacae mutant for bioaugmentation of petroleum- and salt-contaminated soil [J]. Biochemical Engineering Journal, 2010, 49(2): 201-206.
[14] MA Y, YANG H, CHEN X, et al. Significantly improving the yield of recombinant proteins in Bacillus subtilis by a novel powerful mutagenesis tool (ARTP): Alkaline ІС-amylase as a case study [J]. Protein Expr Purif, 2015, 114(82-88.
[15] FAN X, WU H, LI G, et al. Improvement of uridine production of Bacillus subtilis by atmospheric and room temperature plasma mutagenesis and high-throughput screening [J]. Plos One, 2017, 12(5): e0176545.
[16] WANG J X, ZHANG Z Y, LIU H D, et al. Construction and optimization of trans-4-hydroxy-L-proline production recombinant E-coli strain taking the glycerol as carbon source [J]. Journal of Chemical Technology and Biotechnology, 2016, 91(9): 2389-2398.
[17] LIU R M, LIANG L Y, MA J F, et al. An engineering Escherichia coli mutant with high succinic acid production in the defined medium obtained by the atmospheric and room temperature plasma [J]. Process Biochem, 2013, 48(11): 1603-1609.
[18] WANG Y, LI Q G, ZHENG P, et al. Evolving the L-lysine high-producing strain of Escherichia coli using a newly developed high-throughput screening method [J]. Journal of Industrial Microbiology & Biotechnology, 2016, 43(9): 1227-1235.
[19] LI H P, WANG Z B, GE N, et al. Studies on the Physical Characteristics of the Radio-Frequency Atmospheric-Pressure Glow Discharge Plasmas for the Genome Mutation of Methylosinus trichosporium [J]. Ieee Transactions on Plasma Science, 2012, 40(11): 2853-2860.
[20] DONG T T, GONG J S, GU B C, et al. Significantly enhanced substrate tolerance of Pseudomonas putida nitrilase via atmospheric and room temperature plasma and cell immobilization [J]. Bioresource Technology, 2017, 244(1104-1110.
[21] SHI F, TAN J, CHU J, et al. A qualitative and quantitative high-throughput assay for screening of gluconate high-yield strains by Aspergillus niger [J]. Journal of Microbiological Methods, 2015, 109(134-139.
[22] ZHU X, ARMAN B, CHU J, et al. Development of a method for efficient cost-effective screening of Aspergillus niger mutants having increased production of glucoamylase [J]. Biotechnology Letters, 2017, 39(5): 1-6.
[23] WANG Q, FENG L R, WEI L, et al. Mutation Breeding of Lycopene-Producing Strain Blakeslea Trispora by a Novel Atmospheric and Room Temperature Plasma (ARTP) [J]. Applied Biochemistry and Biotechnology, 2014, 174(1): 452-460.
[24] QIN T T, SONG P, WANG X T, et al. Protoplast mutant selection of Glarea Lozoyensis and statistical optimization of medium for pneumocandin B-0 yield-up [J]. Bioscience Biotechnology and Biochemistry, 2016, 80(11): 2241-2246.
[25] LI X Y, LIU R J, LI J, et al. Enhanced arachidonic acid production from Mortierella alpina combining atmospheric and room temperature plasma (ARTP) and diethyl sulfate treatments [J]. Bioresource Technology, 2015, 177(134-140.
[26] HAIMOVICH A D, MUIR P, ISAACS F J. Genomes by design [J]. Nature Reviews Genetics, 2015, 16(9): 501-516.
[27] LIN J L, WAGNER J M, ALPER H S. Enabling tools for high-throughput detection of metabolites: Metabolic engineering and directed evolution applications [J]. Biotechnology Advances, 2017, 35(8): 950-970.
[28] XIA Q, ZHANG H, SUN X, et al. A comprehensive review of the structure elucidation and biological activity of triterpenoids from Ganoderma spp [J]. Molecules, 2014, 19(11): 17478-17535.
[29] XU J W, ZHAO W, ZHONG J J. Biotechnological production and application of ganoderic acids [J]. Applied Microbiology & Biotechnology, 2010, 87(2): 457.
[30] СѕОЉЛЊ. ЛљгкЩњЮяЙтЦзЖдЕШРызгЬхгеБфгъЩњКьЧђдхЛњРэМАЭЛБфЬхПьЫйЩИбЁЗНЗЈЕФбаОП [D]; жаЙњПЦбЇММЪѕДѓбЇ, 2016.
[31] ЛЦЧрЃЌЙтЦздкЩњЮябЇбаОПжаЕФгІгУ [J]. АВЛеДѓбЇбЇБЈЃЈздШЛПЦбЇАцЃЉЃЌ2012ЃЌ36ЃЈ1ЃЉЃК6-17.
[32] СѕОЉЛЊЃЌЛЦЧр. FT-IR ЙтЦздкЕчРыЗјЩфзїгУгкЮЂЩњЮябаОПжаЕФгІгУ [J].ЩњЮяЮяРэбЇБЈЃЌ2012ЃЌ28ЃЈ9ЃЉ:1-12.
[33] грдіСС, ЭѕМЭ, дЌГЩСш, ЕШ. ЮЂЩњЮягЭжЌЛЈЩњЫФЯЉЫсВњЩњОњРызгЪјгеБфКЭЗЂНЭЕїПи [J]. ПЦбЇЭЈБЈЃЌ2012ЃЌ57ЃЈ11ЃЉЃК883-890.
[34] SANTOS C, FRAGA M E, KOZAKIEWICZ Z, et al. Fourier transform infrared as a powerful technique for the identification and characterization of filamentous fungi and yeasts [J]. Research in Microbiology, 2010, 161(2): 168-175.
[35] FISCHER G, BRAUN S, THISSEN R, et al. FT-IR spectroscopy as a tool for rapid identification and intra-species characterization of airborne filamentous fungi [J]. Journal of Microbiological Methods, 2006, 64(1): 63-77.
[36] BURATTINI E, CAVAGNA M, DELLЁЏANNA R, et al. A FTIR microspectroscopy study of autolysis in cells of the wine yeast Saccharomyces cerevisiae [J]. Vibrational Spectroscopy, 2008, 47(2): 139-147.
[37] CORTE L, RELLINI P, ROSCINI L, et al. Development of a novel, FTIR (Fourier transform infrared spectroscopy) based, yeast bioassay for toxicity testing and stress response study [J]. Analytica Chimica Acta, 2010, 659(1ЈC2): 258-265.
[38] HOLMAN H Y, WOZEI E, LIN Z, et al. Real-time molecular monitoring of chemical environment in obligate anaerobes during oxygen adaptive response [J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(31): 12599.
[39] MELIN A M, PERROMAT A, DELERIS G. Sensitivity of Deinococcus radiodurans to ІУ-Irradiation: A Novel Approach by Fourier Transform Infrared Spectroscopy [J]. Archives of Biochemistry & Biophysics, 2001, 394(2): 265-274.
[40] СѕОЉЛЊЃЌЛЦЧр. гІгУКьЭтЙтЦзЯдЮЂГЩЯёММЪѕЙлВтЕЅЯИАћгъЩњКьЧђдхЯКЧрЫиДњаЛЙ§ГЬ [J].ЙтЩЂЩфбЇБЈЃЌ2014ЃЌ,26ЃЈ3ЃЉЃК321-325.
[41] LIU J, CHEN J, CHEN Z, et al. Isolation and characterization of astaxanthin-hyperproducing mutants of Haematococcus pluvialis (Chlorophyceae) produced by dielectric barrier discharge plasma [J]. Phycologia, 2016, 55(6): 650-658.
[42] LIU J, HUANG Q. Screening of Astaxanthin-Hyperproducing Haematococcus pluvialis Using Fourier Transform Infrared (FT-IR) and Raman Microspectroscopy [J]. Applied Spectroscopy, 2016, 70(
[43] LIU J H, SONG L, HUANG Q. Rapid screening astaxanthin-hyperproducing Haematococcus pluvialis mutants through near infrared spectroscopy [J]. Letters in Applied Microbiology, 2016, 62(2): 185-191.
[44] СѕОЉЛЊ, ГТОќ, ЧиЫЩЃЌЕШ. КьЭтЙтЦздкЮЂдхСьгђЕФгІгУбаОПНјеЙ [J]. ЙтЦзбЇгыЙтЦзЗжЮі, 2019, 39(01): 79-86.
[45] ЭѕДПбє, ТэгёК, СѕБѓУРЃЌЕШ. НќКьЭтЭИЗДЩфЙтЦзВтЖЈЕЅСЃЕОжжЕФЕААзжЪКЌСП [J]. КЫХЉбЇБЈ, 2019, 33(10): 2003-2012.
[46] MA Y H, ZHANG Q Q, ZHANG Q F, et al. Improved production of polysaccharides in Ganoderma lingzhi mycelia by plasma mutagenesis and rapid screening of mutated strains through infrared spectroscopy [J]. Plos One, 2018, 13(9):
[47] SKALICKA-WOZNIAK K, SZYPOWSKI J, LOS R, et al. Evaluation of polysaccharides content in fruit bodies and their antimicrobial activity of four Ganoderma lucidum (W Curt.: Fr.) P. Karst. strains cultivated on different wood type substrates [J]. Acta Societatis Botanicorum Poloniae, 2012, 81(1): 17-21.
[48] MA Y, HE H, WU J, et al. Assessment of Polysaccharides from Mycelia of genus Ganoderma by Mid-Infrared and Near-Infrared Spectroscopy [J]. Scientific Reports, 2018, 8(1): 1-10.
[49] NAUMANN A, NAVARRO-GONZALEZ M, PEDDIREDDI S, et al. Fourier transform infrared microscopy and imaging: detection of fungi in wood [J]. Fungal Genet Biol, 2005, 42(10): 829-835.
[50] KOZARSKI M, KLAUS A, NIKSIC M, et al. Antioxidative and immunomodulating activities of polysaccharide extracts of the medicinal mushrooms Agaricus bisporus, Agaricus brasiliensis, Ganoderma lucidum and Phellinus linteus [J]. Food Chemistry, 2011, 129(4): 1667-1675.
[51] GHEBRESELASSIE H. Measuring the hydrophobicity of cellulose and the effect of humidity by inverse gas chromatography [J]. Dissertations & Theses - Gradworks, 2013,
[52] TANGSADTHAKUN C, KANOKPANONT S, SANCHAVANAKIT N, et al. The influence of molecular weight of chitosan on the physical and biological properties of collagen/chitosan scaffolds [J]. J Biomater Sci Polym Ed, 2007, 18(2): 147-163.
[53] CHEN X, LIU X, SHENG D, et al. Distinction of broken cellular wall Ganoderma lucidum spores and G. lucidum spores using FTIR microspectroscopy [J]. Spectrochim Acta A Mol Biomol Spectrosc, 2012, 97(667-672.
[54] CARRASCO H, ESPINOZA L, CARDILE V, et al. Eugenol and its synthetic analogues inhibit cell growth of human cancer cells (part I) [J]. Journal of the Brazilian Chemical Society, 2008, 19(3): 543-548.
[55] LIU G, SONG D S, ZHAO D Z, et al. A study of the mushrooms of boletes by Fourier transform infrared spectroscopy [M]//BALLY G V, LUO Q. Ico20: Biomedical Optics. 2006.
[56] MURESAN A E, MUSTE S, VLAIC R A, et al. HPLC Determination and FT-MIR Prediction of Sugars from Juices of Different Apple Cuhivars during Fruit Development [J]. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 2015, 43(1): 222-228.
[57] OJAMAE L, HERMANSSON K, PROBST M. THE OH STRETCHING FREQUENCY IN LIQUID WATER SIMULATIONS - THE CLASSICAL ERROR [J]. Chemical Physics Letters, 1992, 191(5): 500-506.
[58] ZHANG F, HUANG Q, YAN J, et al. Assessment of the Effect of Trichostatin A on He La Cells through FT-IR Spectroscopy [J]. Analytical Chemistry, 2015, 87(4): 2511-2517.
[59] XUE Q, PENG W X, OHKOSHI M. Molecular bonding characteristics of Self-plasticized bamboo composites [J]. Pakistan Journal of Pharmaceutical Sciences, 2014, 27(4): 975-982.
[60] ZHU X L, CHEN A F, LIN Z B. Ganoderma lucidum polysaccharides enhance the function of immunological effector cells in immunosuppressed mice [J]. Journal of Ethnopharmacology, 2007, 111(2): 219-226.
[61] GOO B G, BAEK G, CHOI D J, et al. Characterization of a renewable extracellular polysaccharide from defatted microalgae Dunaliella tertiolecta [J]. Bioresource Technology, 2013, 129(343-350.
[62] SHAO L, WU Z J, ZHANG H, et al. Partial characterization and immunostimulatory activity of exopolysaccharides from Lactobacillus rhamnosus KF5 [J]. Carbohydrate Polymers, 2014, 107(51-56.
[63] ZHAO T, MAO G H, FENG W W, et al. Isolation, characterization and antioxidant activity of polysaccharide from Schisandra sphenanthera [J]. Carbohydrate Polymers, 2014, 105(26-33.
[64] GRUBER J, LI R W C, PONCE P, et al. Novel soluble blue emitting PPV-like polymers: synthesis and characterization [J]. E-Polymers, 2003,
[65] ETCHEVERRY S B, WILLIAMS P A M, BARAN E J. Synthesis and characterization of oxovanadium(IV) complexes with saccharides [J]. Carbohydrate Research, 1997, 302(3-4): 131-138.
[66] PETROSYAN A M. Vibrational spectra of L-histidine perchlorate and L-histidine tetrafluoroborate [J]. Vibrational Spectroscopy, 2007, 43(2): 284-289.
[67] DEFREES D J, MILLER M D, TALBI D, et al. THEORETICAL INFRARED-SPECTRA OF SOME MODEL POLYCYCLIC AROMATIC-HYDROCARBONS - EFFECT OF IONIZATION [J]. Astrophysical Journal, 1993, 408(2): 530-538.
[68] CHOONG Y K, SUN S Q, ZHOU Q, et al. Determination of storage stability of the crude extracts of Ganoderma lucidum using FTIR and 2D-IR spectroscopy [J]. Vibrational Spectroscopy, 2011, 57(1): 87-96.
[69] VARETTI E L, AYMONINO P J. Ir Spectra of Perfluoromethyl Perfluoroacetate and Perfluoromethyl Carbonate [J]. Journal of Molecular Structure, 1971, 7(1-2): 155-&.
[70] WORKMAN J, WEYER L. Practical Guide to Interpretive Near-Infrared Spectroscopy [M]. CRC Press, Inc., 2007.
[71] FERREIRA D S, POPPI R J, PALLONE J A L. Evaluation of dietary fiber of Brazilian soybean (Glycine max) using near-infrared spectroscopy and chemometrics [J]. Journal of Cereal Science, 2015, 64(43-47.
[72] FACKLER K, SCHWANNINGER M, GRADINGER C, et al. Qualitative and quantitative changes of beech wood degraded by wood-rotting basidiomycetes monitored by Fourier transform infrared spectroscopic methods and multivariate data analysis [J]. FEMS Microbiol Lett, 2007, 271(2): 162-169.
[73] HE W, HU H. Prediction of hot-water-soluble extractive, pentosan and cellulose content of various wood species using FT-NIR spectroscopy [J]. Bioresource Technology, 2013, 140(299-305.
[74] CHEN Y, DENG J, WANG Y, et al. Study on discrimination of white tea and albino tea based on near-infrared spectroscopy and chemometrics [J]. J Sci Food Agric, 2014, 94(5): 1026-1033.
[75] CHEN J-B, SUN S-Q, ZHOU Q. Data-driven signal-resolving approaches of infrared spectra to explore the macroscopic and microscopic spatial distribution of organic and inorganic compounds in plant [J]. Analytical and Bioanalytical Chemistry, 2015, 407(19): 5695-5706.
[76] CHEN S F, DANAO M G, SINGH V, et al. Determining sucrose and glucose levels in dual-purpose sorghum stalks by Fourier transform near infrared (FT-NIR) spectroscopy [J]. J Sci Food Agric, 2014, 94(12): 2569-2576.
[77] MOHACEK-GROSEV V, BOZAC R, PUPPELS G J. Vibrational spectroscopic characterization of wild growing mushrooms and toadstools [J]. Spectrochim Acta A Mol Biomol Spectrosc, 2001, 57(14): 2815-2829.
[78] He H, Yao G, Ma Y, et al. Experimental and theoretical study of the Raman spectra of ganoderic acid T [J]. J. Structural Chemistry. 2019. 60(9):1467-1480.
[79] YAO G H, MA Y H, MUHAMMAD M, et al. Understanding the infrared and Raman spectra of ganoderic acid A: An experimental and DFT study [J]. Spectrochimica Acta Part a-Molecular and Biomolecular Spectroscopy, 2019, 210(372-380.
[80] еХйЛйЛЃЌЛЦЧр. ЛљгкЯуВнШЉ-ИпТШЫсЯдЩЋЗДгІВтЖЈСщжЅШ§нЦЕФЗНЗЈЬНЬжгыаое§[J]. ОњЮябЇБЈЃЌ2018, 37(12): 1792-1801.
ЩЯвЛЦЊЃКДѓЧђИЧЙНжжжЪзЪдДвХДЋЖрбљадКЭЧздЕЙиЯЕЗжЮі
ЯТвЛЦЊЃКМДЪГаЭТЬВшЫЋцпЙНИёРЬѕЕФбажЦ